|
The evolution of locomotor adaptations
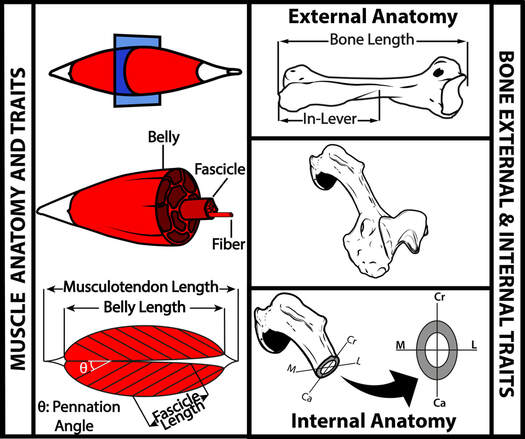
Limbs are comprised of multiple anatomical components, such as muscles and bones. Notably muscles and bones have distinct biomechanical roles, with muscles producing force and bones transmitting it. Moreover, the internal structure of bones determines how a limb is able to stand up to mechanical loads, such as bending and compression. The mutli-component nature of limbs raises the question: if an adaptation occurs in an organ, such as a limb, do all of the organ's components together experience a selective pressure for the adaptation? Mustelid mammals (i.e, weasels, badgers, martens, otters) are the focus of this project, as from a single, common ancestor, the Mustelidae have radiated into species specialized for climbing, digging, and swimming. This makes them an ideal group with which to study the evolution of limb adaptations. This project combines morphological data on limb bones (lengths and diameters, cross-sectional properties, and microanatomy) and limb muscle architecture with phylogenetic data and evolutionary modeling to determine if all the traits that enable limb biomechanical function experience a single set of evolutionary processes. This project is funded by the Deutsche Forschungsgemeinschaft, and results have thus far appeared in Ecology and Evolution, BMC Evolutionary Biology, Royal Society Open Science, and Integrative Organismal Biology. |
|
Biomechanical size and shape in mammalian limbs
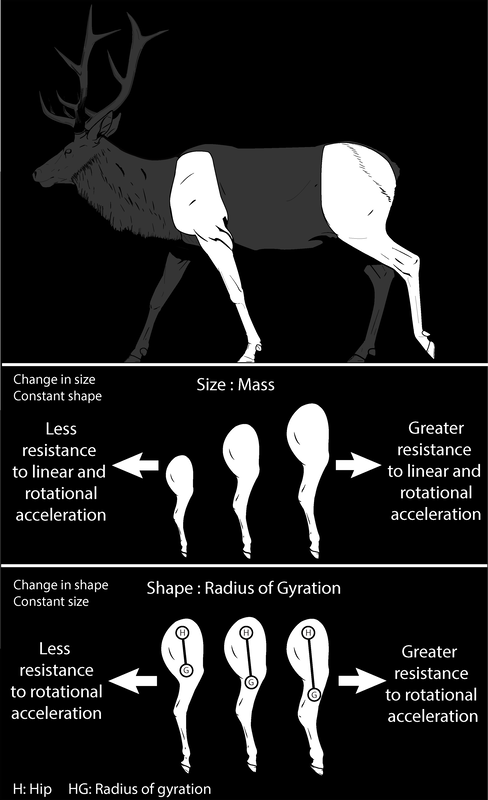
One of the most conspicuous aspects of mammalian diversity is the size and shape of limbs, from the webbed feet of otters to the oversized claws of armadillos, from the svelte limbs of antelope to the heavyset limbs of rhinos. Given that terrestrial locomotion in mammals is the result of the limbs moving back and forth relative to the body, it raises a question: what does this diversity in limb size and shape entail for the biomechanics of terrestrial locomotion? This project gathered measures of limb size and shape for 44 species of mammal, ranging from wolves to wildebeest, from hyrax to zebras. Limb mass was the measure of limb size studied, as mass represents an object’s resistance to linear acceleration, and the radius of gyration was the measure of limb shape, with the radius of gyration reflecting a rotating object’s mass distribution relative its axis of rotation (think of a hindlimb swinging back and forth about the hip). The more mass is concentrated towards the axis of rotation, the shorter the radius of gyration. Limb mass and radius gyration together produce a third variable, the moment of inertia, which in turn is a measure of an object’s resistance to rotational acceleration. My colleagues and I found that, relative to a limb’s length, the mass and moment of inertia are disproportionately lower in larger mammals than smaller mammals, suggesting that in larger limbed ungulates the cost of swinging the limbs may be low relative to their limb’s length. This project was funded by a National Science Foundation Graduate Research Fellowship, the University of Chicago, the American Society of Mammalogists, the Society for Integrative & Comparative Biology, Sigma Xi, the Field Museum of Natural History, and the Wissenschaftskolleg zu Berlin. The results of this study were published in Evolution and PLoS ONE, and related studies have been published in Journal of Experimental Zoology, Frontiers in Zoology, Biological Journal of the Linnean Society, and Journal of Experimental Biology. |
|
Postnatal ontogeny of the locomotor system in mammals and dinosaurs
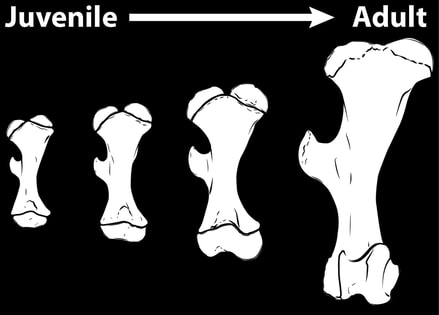
Prior to this project, previous studies suggested that the limb bones of ectothermic and endothermic species exhibited different growth patterns during postnatal ontogeny, as individuals grew from neonates into adults. The limb bones of ectothermic species were suggested to grow more robust or maintain their proportions during postnatal ontogeny, whereas the limb bones of endothermic species were suggested to grow more gracile. However, this hypothesis was based upon only ectothermic species and a handful of smaller-bodied endothermic species. This in turn lead to the question: are the proposed growth patterns distinguishing ectothermic and endothermic species pervasive across a greater sample of species not only in terms of taxonomy but also in terms of size? For this project, my internship (and later Ph.D.) advisor Dr. Peter Makovicky and I compiled a sample of 22 mammal species encompassing six taxonomic orders and a sample of 21 (non-bird) dinosaur species encompassing five major dinosaurs groups. In neither mammals nor dinosaurs did we find a single growth pattern. Instead in mammals overall, species within Carnivora tended to have limb bones that grow more gracile during ontogeny, whereas ungulates tended to have bones that grow more robust during ontogeny. In dinosaurs, limb bones tended to grow more robust during ontogeny or without changing their proportions; however, hadrosaurid (“duck-billed”) dinosaurs tended to have limb bones that grow more gracile during ontogeny. Within mammals, species that have an adult body mass below 15-20 kg tended to have limbs bones that grow increasingly gracile during ontogeny, whereas species that have an adult body mass greater than this range tended to have limb bones that grow with constant proportions or that grow more robust during ontogeny. This project was funded by a National Science Foundation Graduate Research Fellowship and the American Museum of Natural History, and the results were published in Journal of Anatomy and Journal of Morphology. |
